
Article
The Missing Baseline: What Does Healthy Foot Function Actually Look Like?
Introduction
Every clinical field builds its understanding of dysfunction on a definition of what optimal function looks like. Oncology defines cancer against the biology of healthy cells. Cardiology defines disease against the physiology of a healthy heart. Before one can say something is wrong, one needs a clear account of what right looks like.
Conventional foot care has never produced that definition.
For over a century, the profession has studied, categorised, and intervened on foot structure and function — without first establishing what optimal foot function actually requires. What has been observed, measured, and treated as the baseline is a population that has spent its developmental and adult years in conventional footwear. What the profession has called “normal” is the physiological outcome of a specific set of inputs — not the starting point it believes it to be.
That distinction is not semantic. It is the epistemological foundation on which every clinical assessment, every intervention, and every outcome measure in conventional foot care rests. And it has never been examined.
This article examines it.
Section 1: The Historical Error — How Conventional Norms Arose
The conventional foot care profession approaches the foot from two distinct perspectives. The first is anatomical and structural: the foot is examined in terms of its component parts — bone architecture, soft tissue configuration, joint alignment — assessed largely as static entities. Foot type is identified, deviation from an observed norm is measured, and an intervention is prescribed to manage that deviation. The second perspective is gait analysis: dynamic observation of foot and lower limb movement through motion capture, force plates, and kinematic modelling. These appear to represent different domains of enquiry. They do not. Perspective 2 is Perspective 1 fitted with cameras — designed from the structural model and interpreted through its lens.
The most widely used clinical gait analysis tool — the Conventional Gait Model (CGM), implemented in Vicon’s Plug-in Gait system — models the entire foot as a single rigid segment (Davis et al., 1991). The foot’s 26 bones, 33 joints, and the full dynamic interplay between them are collapsed into a single unit. The Oxford Foot Model improved on this with a three-segment approach — hindfoot, forefoot, and hallux tracked separately (Carson et al., 2001) — and most instrumented gait laboratories now integrate force plate kinetics with motion capture kinematics, calculating joint moments and ground reaction forces across the gait cycle. These are genuine technical advances. But even the most fully instrumented configuration cannot capture what defines optimal foot function as a holistic mechanism: the synergistic dynamic rise of the great toe during the swing phase, the elevation of the arch, the formation of the Windlass Mechanism, the stabilizing sesamoid positioning, the cuboid pulley mechanism — events that collectively prepare the feet and ankles before ground contact occurs – the functional mechanics that safely and efficiently manage the mechanical demands of loading throughout the lower limb kinetic chain. Not because this movement is invisible — it is entirely observable on the external surface of the foot to any observer who knows what to look for. But because the measurement protocols were designed from a conceptual model that does not include these events as primary functional architecture, no current system tracks them.
The normative databases built from these systems compound the illusion. Their reference populations are habitually shod subjects — adults and children whose developmental and adult years in conventional footwear have progressively entrained away the capabilities the measurement systems do not capture. Optimal pre-contact functional mechanics during the swing phase — is a neuromuscular capability that requires daily demand to maintain. In a habitually shod population, that demand is largely absent. The capability attenuates. What the normative databases record as typical pre-contact hallux behaviour is the functional output of lifelong restriction. Eggleston et al. (2023) raised this concern explicitly from within the field, cautioning that barefoot walking in habitually shod subjects produces shod-pattern movement without the shoe — and should not be treated as a reference for unencumbered foot function.
The literature’s own canonical description of the primary foot mechanism reinforces the error. The Windlass mechanism — first described by Hicks (1954) and reproduced in the biomechanics literature since — is framed as a reactive, post-contact event: the heel rises, ground reaction forces dorsiflex the toes, and the plantar fascia is tensioned as a consequence.
Wearing et al. (2006) and Wearing (2012) subsequently challenged this characterisation, proposing that the plantar fascia is more accurately understood as a dynamic coordinator of movement — conforming to the contours of the plantar surface rather than spanning it as a passive tension element — and that the arch-raising function attributed to fascial tension may rely more substantially on bone geometry and active muscular mechanisms than the conventional model acknowledges.
Load is the trigger; the mechanism follows. This is a precise description of what occurs in a foot that arrives at ground contact without anticipatory preparation already in place — a reactive compensation for the absence of pre-contact bracing. In optimal function, the sequence is inverted: the great toe dorsiflexes actively during swing, with no load, under voluntary neuromuscular control, the Windlass Mechanism is activated before the loading event begins. A stable dynamic arch system is in place before loading forces are engaged. What Hicks’ widely accepted canonical description of the mechanism does recognise is a description of the attenuated, reactive pattern — observed in habitually shod subjects, published as the mechanism, and taught as such ever since.
Modern epigenetics dismantles the static assumption that underlies both perspectives.
Epigenetic research has established that gene expression is continuously modulated by environmental inputs — including mechanical load, movement demand, sensory stimulus, and physical activity. A single exercise session produces measurable changes in gene expression within hours (Barrès et al., 2012). Lifelong movement patterns produce lasting alterations in how genes are expressed across metabolic, contractile, and structural systems. The body is not a fixed structure that wears over time. It is a dynamic, continuously adapting system whose current state reflects the cumulative instruction it has received through use.
All systems of the body are plastic. They are synergistically integrated and continuously adapting — every moment of every day — to thought, emotion, movement, nutrition, and environment. Nothing is fixed. The direction of adaptation follows the direction of the instruction the body receives.
The foot the profession has been studying is a foot that has received a specific set of instructions — restriction, sensory attenuation, and mechanical unloading — delivered continuously through conventional footwear throughout the developmental and adult years. The functional state that results is not a baseline. It is an outcome. The field’s foundational error was epistemological: it was studying an outcome and calling it a starting point.
Both the maladaptive foot state and what might be considered a healthier foot state are the body’s entirely normal response to the inputs it receives. There is no state that is more or less natural — only the body’s normal adaptive response to different conditions of entrainment. What differs is not the body’s logic, but the instruction it has been given and the functional direction that instruction produces.
Section 2: The Architecture — What the Foot Actually Is
Before evaluating function, it is necessary to understand the structure being evaluated.
The foot and ankle function as a holistic neuromusculoskeletal dynamically synergistic structural platform for the lower limb kinetic chain. This is not a poetic description. It is a precise functional statement — and one that most conventional clinical assessments fail to honour in full.
The foot contains 26 bones, 33 joints, and more than 100 muscles, tendons, and ligaments — organised into a set of interdependent systems, each with a distinct architectural role. Bones provide the skeletal scaffold: the structural geometry through which all mechanical forces pass. Joints — all 33 of them — are the points of articulation that define the axes along which controlled movement between those bones is possible. Ligaments and fascia connect the bones at the joints and govern the permissible range and direction of movement at each articulation, providing the structural continuity that holds the architecture together under load. Cartilage lines the articular surfaces, protecting bone from direct contact and enabling smooth, low-friction movement. Muscles generate movement, through bone-to-bone attachment, and through the tendons that transmit contractile force to more distal structures. The foot’s intrinsic musculature reflects more so bone to bone connections while the foot’s extrinsic muscles have their muscle bodies located in the lower leg, which transmit force into the foot via long tendons.
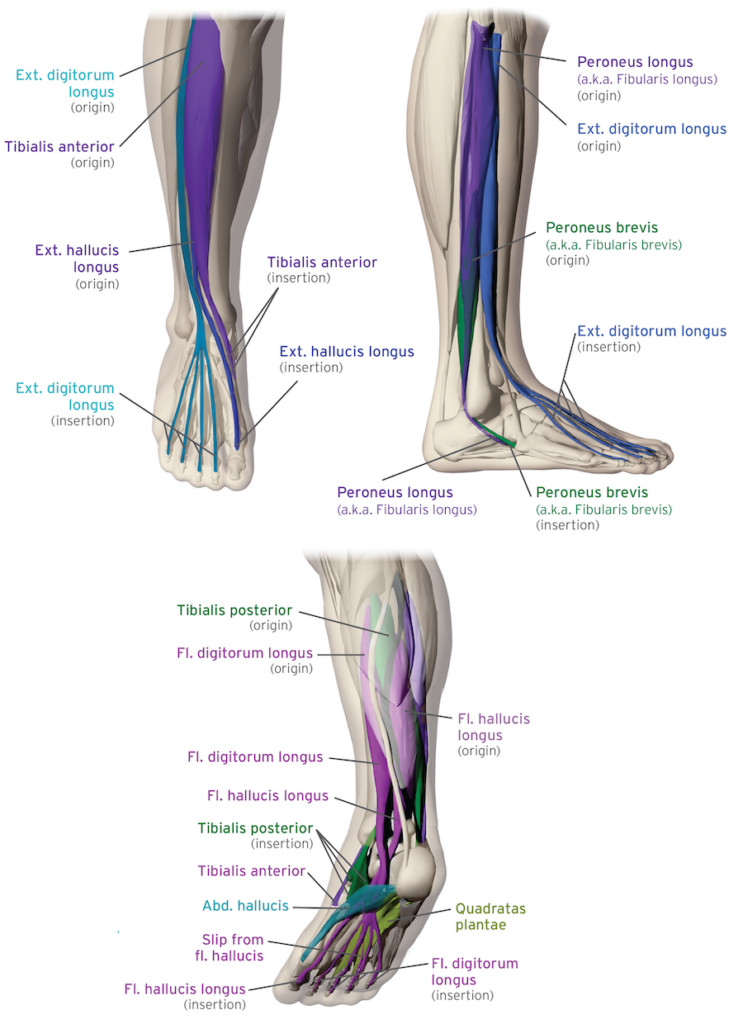
The foot’s extrinsic musculature — each muscle body originates in the lower leg and activates foot movement via long tendons. These are the muscles that drive the pre-contact anticipatory arch preparation sequence described in Section 5.
The vascular system supplies all the above. The arterial network delivers oxygen and nutrients to every tissue in the foot — bone, cartilage, muscle, fascia, and neural structures alike. The venous and lymphatic networks remove metabolic waste and maintain the cleansing processes on which tissue health depends (Arnett, 2013). The foot’s plantar venous plexus contributes a further mechanical function: under correct weight-bearing load, it operates as a peripheral pump — the first stage of venous return during walking, driving blood upward through the deep venous system ahead of the calf pump (Gardner and Fox, 1983; Corley et al., 2010).
These structural numbers — 26 bones, 33 joints, more than 100 muscles, tendons, and ligaments — represent considerable mechanical complexity. They are, however, modest when set against the foot’s sensory infrastructure. The soles of each foot alone contain an estimated 200,000 nerve endings — a figure that accounts for the cutaneous sensory receptors in the plantar skin: Meissner’s corpuscles, Merkel disk receptors, Ruffini endings, and Pacinian corpuscles. It does not include the additional sensory apparatus housed within the plantar fascia itself, which Stecco et al. (2013) established is rich in Pacinian and Ruffini corpuscles and plays a significant proprioceptive role independent of the skin’s contribution. The full sensory density of the sole — skin and fascia combined — is considerably greater than the conventional figure reflects.
The nervous system is the sensory and organisational infrastructure that integrates the entire architecture. Through the foot’s nociceptors, mechanoreceptors, and proprioceptors — distributed across the plantar surface and throughout the medial, lateral, and dorsal aspects of the foot and ankle — it provides the brain with continuous, moment-by-moment information on spatial positioning, loading forces, and the characteristics of the environment the foot is in contact with. The plantar surface, most densely innervated, reads primarily what is beneath the foot. The medial, lateral, and dorsal surfaces read what surrounds it. Together, this sensory web constitutes the primary data stream through which the brain organises movement and assesses the foot’s relationship with its environment.
Each of these systems is distinct in its functional role. None operates independently of the others. How they function as a synergistic whole — and what the laws of physiology require of that whole — is the subject of the section that follows.
Section 3: The Laws of Physiology — What is Actually Happening
All the body’s systems, bone, muscle, fascia, neural, venous, digestive, etc. are plastic; they are constantly adapting – moment to moment – to use and environment.
Every person, regardless of their genetic predisposition, has an achievable functional “sweet spot” – their optimal functional robustness.
The body’s neuromusculoskeletal functional capabilities at any given “time” reflect historical daily usage. Daily activities that promote a balance of strength and flexibility at the joints safely encourages and maintains functional robustness — enhanced capabilities. Daily activities that impose restriction or mitigate sensory inputs entrains functional atrophy.
This is not a theory. It is the convergent conclusion of multiple independent disciplines: Wolff’s Law (bone remodels along lines of habitual mechanical stress), Davis’s Law (soft tissue adapts to demands imposed upon it and atrophies under prolonged support or restriction), the Specific Adaptation to Imposed Demands (SAID) principle, the neuroscience of sensory cortical mapping, and the science of epigenetics. Each of these scientific perspectives make the same fundamental claim from their respective domains: the body becomes what it is consistently asked to do.
Bone adapts to loading forces, muscles adapt and the nervous system re-wires to usage. NASA’s long-duration spaceflight research provides the most controlled evidence for this: astronauts lose 1–1.5% of bone mineral density per month in microgravity — a precise demonstration of Wolff’s Law operating in reverse under conditions of mechanical unloading (Lang et al., 2004; NASA Human Research Program).
Fascia follows the same law. It is not a special case. Fascial integrity is paramount to movement, bodily sense, hormonal, autonomic and neurovascular regulation, and purposeful interaction with our environment (Slater et al., 2024). Fascia is constantly evolving, both dependent on and modulating sustained postures, repetitive movement, quantity of motion, load, stress, strain, hydration, pH, temperature, neurotransmitters, and hormones (Slater et al., 2024).
The fascia’s lubricating medium — hyaluronan — is designed to keep fascial layers sliding freely against one another. Healthy fascia is smooth, slippery and flexible (Johns Hopkins Medicine). Immobility, however, precipitates pathological accumulation of hyaluronan, which super-aggregates in the tissues, impairing blood and lymphatic circulation, and reducing lymphatic efficacy (Slater et al., 2024). Johns Hopkins identifies a lifestyle of limited physical activity — too little movement, day after day — as a primary cause of the fascial adhesion that follows.
The feet and gait-related function are not immune to these adaptive principles – an idea that has most often been overlooked by researchers and healthcare professionals.
The key to understanding what is happening with the foot, is to understand that neuroplasticity plays the leading role.
Most importantly, gait-related neurologic function is a holistic body integrated sensory experience. The foot being an integral system of the functional whole – each system playing a synergistic interdependent role. When the sensory information from one system is attenuated, all systems within the functional whole are directly affected.
Sensory input from the plantar surface of the foot continuously maintains the brain’s cortical representation of foot function in concert with sensory input throughout the kinetic chain. Neural pathways strengthen with use; unused pathways are pruned. The brain reads plantar surface input continuously and uses it to fire protective muscle sequences throughout the kinetic chain ahead of ground contact — a feed-forward system that depends entirely on the quality and variety of afferent input it receives. Attenuate that input and the feed-forward system degrades. The foot arrives at the ground without proficient lower limb alignment in place – such that the kinetic chain is unable to safely and efficiently manage the activity related forces. This attenuated sensory input is uniquely footwear related virtually 99% of the time.
Reduced sensory input from the shod foot — is in fact three compounding mechanisms operating at different levels of the nervous system simultaneously.
The first is mechanical. The shoe sole attenuates ground reaction force magnitude and texture before it reaches the plantar surface. The plantar sensory web reads through a uniform, sensory attenuation filter — and the richness of terrain information the system evolved to process is replaced by a monotonous pressure signature, consistent stride after stride.
The second operates at the peripheral receptor level. The foot’s mechanoreceptors — Meissner’s corpuscles, Merkel disks, Pacinian corpuscles, Ruffini endings — are exquisitely sensitive to change. They are designed to detect variance: texture transitions, edge differentials, load shifts across the plantar surface. Continuous, unchanging input drives these receptors into temporary refractoriness. The sensory web reads uniform pressure as baseline noise rather than as meaningful terrain information. The peripheral filtering compounds the mechanical attenuation before the signal has reached the brain.
The third mechanism operates centrally. Recent neuroscience has identified the orbitofrontal cortex (OFC) as a primary site for long-term predictive filtering of sensory input (Tsukano et al., 2026). The OFC builds an internal model of anticipated sensory experience from accumulated daily exposure. As experience with a consistent sensory environment accumulates, the OFC sends top-down signals to the sensory cortex that actively suppress anticipated input — generating what researchers term a “negative image” of the expected stimulus, cancelling it before conscious processing occurs. In habitually shod feet, the OFC’s predictive model is calibrated to the uniform, attenuated plantar signal that daily footwear delivers. The brain is not passively receiving less information. It is actively cancelling what it has learned to expect.
These three mechanisms compound each other. The shoe attenuates the signal mechanically. The peripheral receptors filter the residual uniform input as noise. The OFC suppresses what it predicts will arrive. The result is a plantar sensory bandwidth far narrower than what the system evolved to operate on — and a feed-forward protective directive generated from that impoverished baseline.
There is a further implication that matters clinically. Because the OFC’s predictive model is built from accumulated experience, removing the shoe does not immediately restore functional efficiency. The predictive suppression persists until the neuromusculoskeletal system is retrained through sustained exposure to varied, rich plantar sensory input. This is the neurobiological mechanism underlying the observation Eggleston et al. (2023) documented: habitually shod subjects walking barefoot produce shod-pattern movement without the shoe. The sensory attenuation has not been reversed by removing its source. The brain continues to operate from the model it built.
It’s not just support and/or restrictiveness that entrain maladaptive function, artificial cushioning entrains maladaptive function as well. Any time, any part of the physiological system is not challenged to do “its job”, that part is being entrained to dysfunction and functional atrophy. The structures that manage mechanical load — bone architecture, fascial web, intrinsic and extrinsic musculature — require differentiating loading to maintain their functional integrity. Remove it, and they adapt accordingly. This is not pathology. It is the body’s entirely normal and predictable response to the sensory input it receives.
Therefore, regardless of genetic predisposition, at any given time, every person’s unique foot and lower limb, hip, and back functional alignment, flexibility, and movement is predicated on multi-system sensory input, or the lack thereof.
Many clinical protocols recommend stretching as the remedy for the resulting stiffness and restriction. McGill University’s Office for Science and Society, reviewing the scientific literature on static stretching, found its track record “unimpressive” — with 119 performance measures showing stretching made performance significantly worse, against only 6 where it improved (Behm et al., 2016; Jarry, McGill OSS, 2022). Afonso et al. found that strength training through full range of motion produced equivalent improvements in joint range of motion to stretching — while additionally developing strength. Prescribing post-workout stretching routines, the researchers concluded, is “based on belief, not data.” Harvard Health confirms this observation: stretching unprepared tissue can damage it — “when everything is cold, the fibers aren’t prepared and may be damaged.”
What the evidence actually supports, is daily dynamic activity that promotes a balance of strength and flexibility at the joints — movement that invites functional adaptation rather than forcing tissue toward a range it may not be prepared to manage. For the past 20 years, this is the primary recommendation of every evidence-based rehabilitation and modern sports training protocol. It is the principle that governs joint and muscle health throughout the body.
The overriding question is: Why do footcare professionals overlook these fundamental principles when viewing the foot and gait-related function?
Section 4: The Elephant in the Room
No published scientific or academic literature defines the mechanics of ideal or optimal gait.
This is not a peripheral gap in the research. It is a foundational absence at the epistemological centre of conventional foot-related hypotheses that frame foot care treatment methodologies and footwear design.
Clinical assessments of foot shape and function measure deviation from an observed population norm. That norm is derived from populations whose feet have spent the developmental and adult years in conventional footwear — populations already subject to the functional entrainment described in Section 3. Conventional foot care has been, in effect, measuring the distance between two adapted states and calling those measurements the normal anatomical shape and the range of normal function.
Johns Hopkins Medicine advises that people with sedentary occupations should take at least a two-minute break every hour to stand up and move around, because movement helps fascia stay supple. The advice is physiologically sound. The variable it does not address is that the person doing the standing and walking does so in footwear that continues to deliver restriction, sensory attenuation, and mechanical unloading to the fascial system with every step. The recommendation is correct. The context in which it is delivered undermines it.
Without a definition of optimal foot function, dysfunction cannot be defined with precision. One cannot measure the distance between where something is and where it should be if the destination has never been specified. The conventional approach has produced a century of interventions aimed at managing deviation from a norm that is itself a product of prior intervention.
This is the perspective conventional foot care often blindly overlooks. Before one can understand how foot problems arise and how best to address them, one needs to understand what optimal function actually looks like.
Section 5: What Optimal Lower Extremity Function Looks Like — Through the Eyes of the Laws of Physiology
Movement capabilities are not genetically “fixed”; they are an entrained response to usage — regardless of genetic predisposition.
The nervous system does not initiate movement from a fixed template. It initiates the best available movement solution given the sensory information it has at any given moment. When sensory information is rich, varied, and unattenuated, the nervous system reflexively initiates the safest, most mechanically efficient, and protective movement possible — optimal neuromusculoskeletal function. Over time, this optimal function becomes the entrained “functional norm”.
When sensory input is attenuated or movement constrained, the nervous system cascades to the next available option: the safest and most energy-efficient solution available within the imposed limits — compensatory neuromusculoskeletal function. Over time, this compensatory function becomes the entrained “dysfunctional norm”.
This is not a compensatory failure. It is the system working exactly as defined by the laws of physiology.
The nervous system’s safety-first reflexive mechanisms take two forms: anticipatory and reactive. Anticipatory reflexes: when we trip and reflexively reach out, arms extended, to break the fall; or duck before a thrown object reaches us. Reactive reflexes: blinking when something enters the eye; coughing to clear the throat; pulling a hand away when it touches something hot.
In gait, anticipatory reflex responses occur throughout the kinetic chain during the swing phase, prior to each foot contacting the ground. The related muscle activations optimally align the structure for safe, efficient, stress-free movement while the feet are on the ground.
Reactive reflex responses occur throughout the kinetic chain while the feet are on the ground. These activations complement the anticipatory alignment — or compensate for its absence.
Franklin and Wolpert (2011) provided the mechanistic basis for the anticipatory side of this picture: the brain builds predictive forward models from accumulated sensory experience to generate feed-forward movement solutions; attenuate that sensory experience, and the anticipatory solutions available to the nervous system are constrained accordingly.
Horak and Nashner (1986) documented the adaptive/reactive cascade that follows in postural control, identifying a hierarchy of movement strategies — ankle, hip, and stepping — that the nervous system selects based on available sensory information and the magnitude of perturbation. The ankle strategy, which requires the most precise sensory input and produces the most mechanically efficient response, is the preferred solution. When sensory input is insufficient, the nervous system cascades to the hip strategy, then to the stepping strategy. It does not fail — it selects the next best available option. Todorov and Jordan (2002), formalising this in the theory of optimal feedback control, demonstrated that the nervous system minimises a composite cost function — task success, energy expenditure, and mechanical safety — selecting from available movement solutions accordingly.
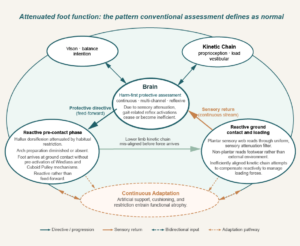
Taken together, these three independent frameworks reach the same conclusion: the movement pattern the nervous system produces is always the optimal solution within its current constraints. Change the constraints, and the solution changes. The three-layer attenuation mechanism described in Section 3 is precisely such a constraint — narrowing plantar sensory bandwidth and forcing the nervous system down the cascade. What the diagram Attenuated foot function: the pattern conventional assessment defines as normal represents is not a dysfunctional foot. It is the nervous system selecting the safest available compensatory pattern within the limits imposed by lifelong footwear-related sensory attenuation.
This brings us to the obvious question: what musculoskeletal movement does the nervous system initiate when none of those constraints are operating? When plantar sensory bandwidth is full, receiving rich, varied terrain information across both the cutaneous and fascial networks, when the OFC’s predictive model is calibrated to unfiltered ground contact? When the lower limb is not artificially braced or its movement artificially restricted? The answer is not a theory. It is realized when gait mechanics is observed through the eyes of the laws of physiology.
When viewed through the eyes of the laws of physiology, an overview of the mechanics of gait suggests a fundamental simplicity: Each step’s lower limb kinetic chain functional mechanics is predicated on the sensory information received during the preceding step along with movement intent. Between each step’s ground contact the neurologic system uses this sensory information to anticipate how to most safely and efficiently manage the oncoming loading forces given the terrain.
During the swing phase of gait, prior to foot strike, anticipatory reflexes activate the lower limb kinetic chain muscles to prepare the chain for the forces generated at and during ground contact. The effectiveness of this synergistic muscle activity is predicated on each component within the kinetic chain fulfilling its respective role.
Since the foot provides the foundation for what occurs mechanically above it, its functional capabilities are paramount for safe, efficient kinetic chain movement.
The foot and ankle’s ability to safely and actively manage a wide range of activity-related forces is predicated on the pre-ground contact synergistic co-activation of the lower leg’s extensor and fibularis system. The extensor hallucis longus raises the great toe, creating the Windlass Mechanism. The tibialis anterior elevates the medial arch while the fibularis longus tendon creates an opposing transverse tension through the cuboid bone (the Cuboid Pulley effect) — cinching the midfoot and raising the lateral arch simultaneously, in concert with the fibularis brevis. These muscles along with the extensor digitorum longus fire together, cinching the calcaneus, cuboid, navicular, cuneiform, and metatarsal bones to form an integrated three-dimensional dynamic dome-like arch system under load. The tibialis posterior co-activates to further stabilize the ankle. Collectively, this pre-ground contact cinching action creates an opposing tension in the extrinsic flexors, further stabilizing the dome-like arch system and ankle throughout weight-bearing from heel strike to toe-off and propulsion.
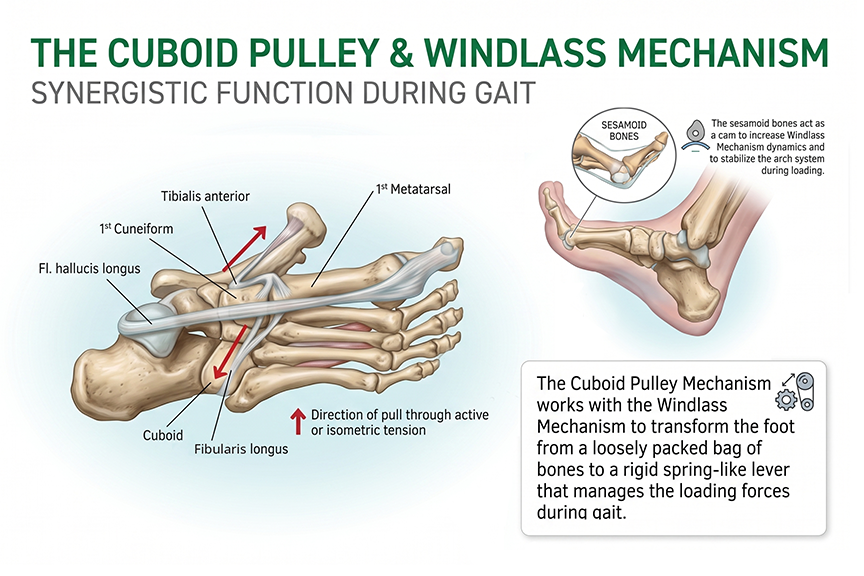
This dome-arch system dynamically distributes vertical forces globally across the structure rather than concentrating stress at any single point. The higher the dome apex, the greater the load management capacity — and the lower the tension imposed on the plantar fascial web — this is simple physics. The Optimal Arch Apex is therefore not a static ideal but is defined by the system’s ability to dynamically adapt and efficiently manage activity-related forces — ensuring that no destructive tension is placed on the fascial web.
The key to the mechanical efficiency and functional strength of this dome-like arch system is the unrestricted ability of the great toe and arch to rise freely in anticipation of loading forces.
The great toe and sesamoid bones contribute a further stabilizing role to this pre-ground contact sequence. As the great toe dorsiflexes, the sesamoids embedded in the flexor hallucis longus move forward and up around the first metatarsal head, increasing the tension — amplifying the Windlass Mechanism. When the first metatarsal head is loaded during weight-bearing, the sesamoids become “trapped” in their forward position. This further inhibits arch collapse or deflection while facilitating the foot’s smooth rotation around the forefoot at toe-off — optimizing propulsion.
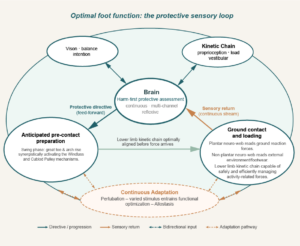
This is the sequence depicted in Optimal foot function: the protective sensory loop — the full anticipatory cascade, operating as the laws of physiology define it.
This dynamic dome-like arch system must be initiated before ground contact, not after it. Optimal gait is anticipatory — feed-forward, not reactive. Why? Because the smaller extrinsic muscles that initiate the pre-ground contact arch system alignment are not strong enough to effectively stabilize — raise the arches as mechanically required — to safely and efficiently manage increased loading forces. The small intrinsic foot muscles cannot efficiently manage these increasing loads as well. When anticipatory activation is absent, the nervous system initiates reactive compensatory muscle activity that is considerably less efficient at safely managing gait-related forces.
This pre-ground contact feed-forward reflex is the nervous system operating at the top of the strategy hierarchy: selecting the most mechanically efficient solution because the sensory information to initiate it is available. This ensures that the feet provide a stable and efficient foundation for the weight-bearing kinetic chain during all gait-related activities.
Every phase of gait is influenced by the one before it. Every phase reflects the functional integrity of the neuromuscular system that drives it — a system that produces this sequence only when the sensory input that sustains it is available, unattenuated, and rich enough to support the optimal solution rather than the compensatory one.
Section 6: The Logical Conclusion
The body becomes what it is asked to do. This is not a philosophical position. It is the convergent conclusion of epigenetics, neuroplasticity, connective tissue physiology, and bone remodelling science — independently derived, consistently confirmed.
Therefore, the foot’s functional capacity at any moment is a precise reflection of the history of demands made of it. A history of dynamic, varied, progressively challenging activity entrains optimal foot function — the capability of safely managing activity-related load and the sensory acuity that the brain requires to anticipate it. A history of restriction, sensory attenuation, and artificial support and cushioning entrains the opposite — not because the body has failed, but because it has succeeded perfectly in adapting to exactly the instruction it received.
What the neuroscience now confirms is that the attenuation does not stop at the periphery. The orbitofrontal cortex builds a predictive model from accumulated sensory experience, actively suppressing anticipated plantar input through top-down inhibitory signalling. The brain is not passively receiving less. It is actively cancelling what it has learned to expect. This distinction has a direct clinical implication: removing the shoe does not restore the system. The predictive model persists. Restoration requires active retraining of the central model — not simply re-exposure to the stimulus.
This is why the cascade principle is not merely descriptive. When sensory input is impoverished, the nervous system does not fail — it reorganizes. It selects the safest available strategy from a diminished set of options, and that selection becomes the new entrained pattern. The dysfunctional norm is not pathology. It is the nervous system operating exactly as designed — optimising for the conditions it has been given.
We now have a definition of optimal function — grounded in the Laws of Physiology, in the fascial and neuromusculoskeletal architecture of the foot, and in what the sensory system requires to initiate the anticipatory cascade across all phases of gait. Measured against that definition, the question shifts. The foot that a century of clinical practice has studied, treated, and called normal is already the precise outcome of prior entrainment. What, precisely, has been measured? And what have a century of interventions been aiming toward?
These are not rhetorical questions. They are the starting point for a different kind of inquiry — one grounded in what the Laws of Physiology actually define, in what the fascial and neuromusculoskeletal systems are actually capable of, and in what optimal functional mechanics actually looks like across all phases of gait.
The practical implications reach further still. If the foot the profession has been studying, treating, and designing footwear around is itself the outcome of lifelong shoe-related entrainment, then many of the pathologies it has catalogued — plantar fasciitis, hallux valgus, metatarsalgia, chronic ankle instability, and the broader catalogue of lower limb overuse conditions — are more accurately described as shoe-related pathologies than foot-related ones. And what optimal treatment methodologies and rehabilitation therapies look like, grounded in a genuine definition of optimal function, is a question the profession has not yet been in a position to ask.
That inquiry — into what is possible, and what the future of foot care and footwear design looks like — starts here.
Sources
Davis RB, Ounpuu S, Tyburski D, Gage JR. A gait analysis data collection and reduction technique. Human Movement Science. 1991;10(5):575–587.
Carson MC, Harrington ME, Thompson N, O’Connor JJ, Theologis TN. Kinematic analysis of a multi-segment foot model for research and clinical applications: a repeatability analysis. Journal of Biomechanics. 2001;34(10):1299–1307.
Eggleston JD, Conroy KE, Moreno AG, Travis WJ, Huskey BR, Vanderhoof HR. The use of preferred footwear versus barefoot conditions in gait analysis: A methodological investigation. Journal of Biomechanics. 2023;160:111817.
Barrès R, Yan J, Egan B, et al. Acute exercise remodels promoter methylation in human skeletal muscle. Cell Metabolism. 2012;15(3):405–411.
Hicks JH. The mechanics of the foot. II. The plantar aponeurosis and the arch. Journal of Anatomy. 1954;88(1):25–30.
Wearing SC, Smeathers JE, Urry SR, Hennig EM, Hills AP. The pathomechanics of plantar fasciitis. Sports Medicine. 2006;36(7):585–611.
Wearing SC. Anatomy of the plantar fascia. In: Schleip R, Findley TW, Chaitow L, Huijing PA, eds. Fascia: The Tensional Network of the Human Body. Churchill Livingstone/Elsevier; 2012. Chapter 5.9.
Slater AM, Barclay SJ, Granfar RMS, Pratt RL. Fascia as a regulatory system in health and disease. Frontiers in Neurology. 2024;15:1458385.
Johns Hopkins Medicine. Muscle Pain: It May Actually Be Your Fascia. hopkinsmedicine.org
Jarry J. Do We Really Need to Stretch? McGill University Office for Science and Society. March 2022.
Harvard Health Publishing. The Importance of Stretching. health.harvard.edu
Behm DG, Blazevich AJ, Kay AD, McHugh M. Acute effects of muscle stretching on physical performance, range of motion, and injury incidence in healthy active individuals: a systematic review. Applied Physiology, Nutrition, and Metabolism. 2016;41(1):1–11.
Afonso J et al. Strength Training versus Stretching for Improving Range of Motion: A Systematic Review and Meta-Analysis. Healthcare. 2021;9(4):427.
Horak FB, Nashner LM. Central programming of postural movements: adaptation to altered support-surface configurations. Journal of Neurophysiology. 1986;55(6):1369–1381.
Todorov E, Jordan MI. Optimal feedback control as a theory of motor coordination. Nature Neuroscience. 2002;5(11):1226–1235.
Franklin DW, Wolpert DM. Computational mechanisms of sensorimotor control. Neuron. 2011;72(3):425–442.
Gardiner R, Comstock B, Dubé S. Biopods Monograph. Biopods Inc.
Tsukano H, Garcia MM, Dandu PR, Kato HK. Orbitofrontal cortex drives predictive filtering of sensory responses. Nature Neuroscience. 2026;29:888–900.
Stecco C, Corradin M, Macchi V, Morra A, Porzionato A, Biz C, De Caro R. Plantar fascia anatomy and its relationship with Achilles tendon and paratenon. Journal of Anatomy. 2013;223(6):665–676.
Lang T, LeBlanc A, Evans H, Lu Y, Genant H, Yu A. Cortical and trabecular bone mineral loss from the spine and hip in long-duration spaceflight. Journal of Bone and Mineral Research. 2004;19(6):1006–1012.
NASA Human Research Program. Risk of Spaceflight-Induced Bone Changes. nasa.gov
Gardner AMN, Fox RH. The venous foot pump: influence on tissue perfusion and prevention of venous thrombosis. Annals of the Royal College of Surgeons of England. 1983;65(2):119–123.
Corley JM, Cross KJ, Hill MK, Gardner AMN. The Anatomy and Physiology of the Venous Foot Pump. The Anatomical Record. 2010;293(5):1–10.
Arnett TR. The Key Role of the Blood Supply to Bone. Bone Research. 2013;1(3):203–215.
Article 1 of the Barefootscience.ai content series. 2026-05-29.
Educational content only. This article does not diagnose, treat, or provide personal medical advice. People with pain, diabetes, neuropathy, severe deformity, recent surgery, or other clinical concerns should consult a qualified health professional.